Glycolysis is the biochemical pathway that strips glucose of its energy. It starts with glucose and ends with pyruvate. Pyruvate is converted to acetyl-CoA, which can be stored as fat, or further metabolized in the citric acid cycle. The fate of pyruvate depends on the energy needs of the cell.
The net yield of energy from glycolysis is two adenosine triphosphates (ATP) and two nicotinamide adenine dinucleotide (NADH) molecules per glucose "burned" by the pathway. Each NADH molecule from glycolysis eventually generates 1.5 ATPs from the electron transport chain. Therefore, a total of 5 ATP molecules are produced per molecule of glucose consumed by the glycolysis pathway.
The net energy used or created (in the form of ATP or NADH) are shown in the schematic below:

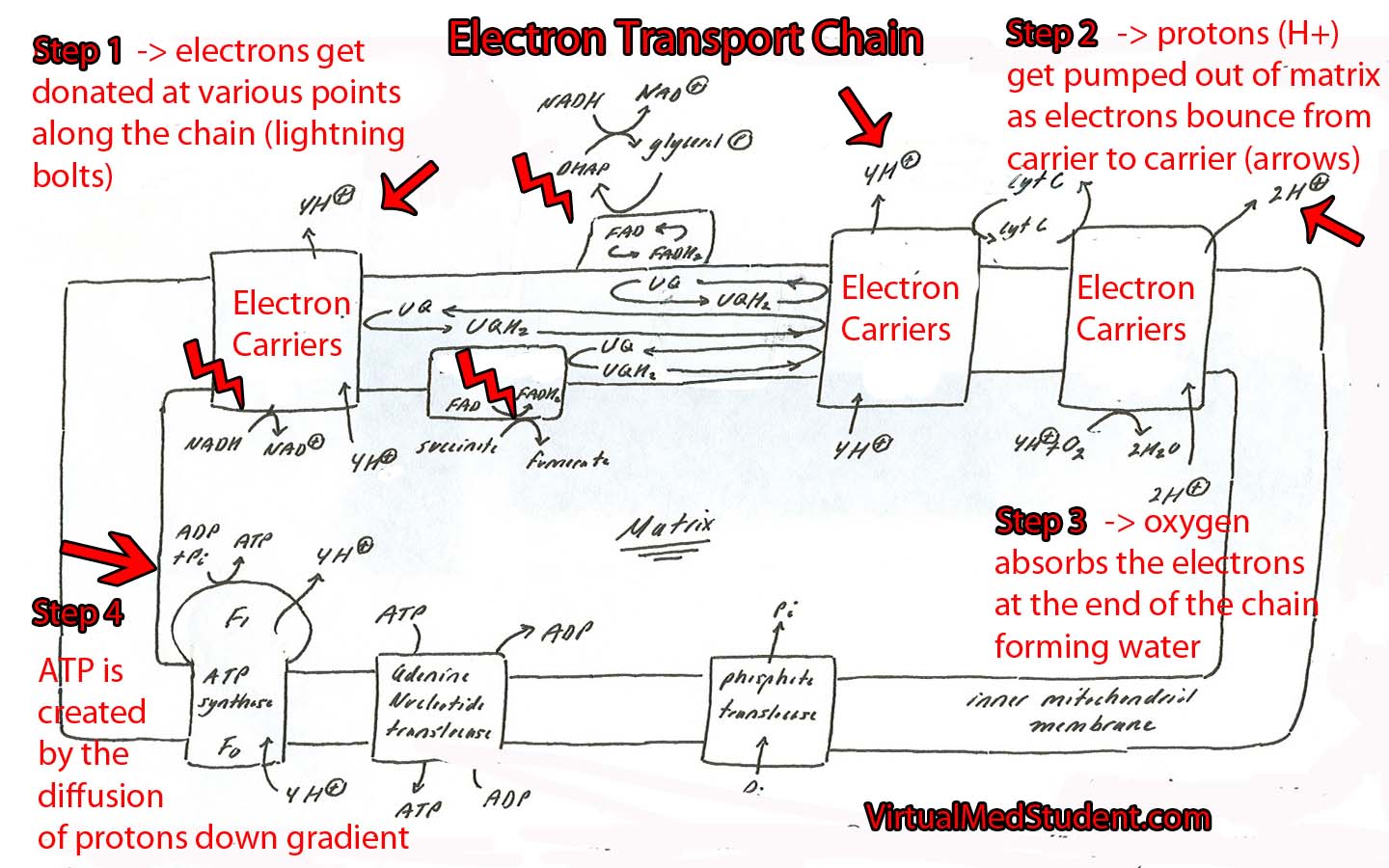
Want to know what happens to the NADH? Check out the article on the electron transport chain.
Regulation
Glycolysis is regulated at several steps in order to ensure that glucose molecules are used appropriately by the cell. The first point of regulation is at the conversion of glucose to glucose-6-phosphate (G6P). This reaction is catalyzed by an enzyme known as hexokinase. This enzyme is inhibited by its own product – G6P. This is known as "feedback inhibition". Once glucose is converted to G6P it becomes "trapped" inside the cell. Therefore, when there are adequate G6P levels any particular cell can shut off the flow of glucose into glycolysis so that it can be sent to other cells that may need it.
(PFK1) is the key
regulatory enzyme in the
glycolytic pathway.
Side note: fructose-2,6-bisphosphate is produced by the enzyme phosphofructokinase-2 (PFK2) and degraded by the enzyme fructose bisphosphatase-2 (FBPase2). PFK2 becomes active under conditions of satiety (ie: well fed states in which lots of glucose/sugar is being absorbed from the gut). When PFK2 is active the concentration of fructose-2,6-bisphosphate increases. This activates PFK1 and increases the activity of the glycolytic pathway.
The final point of regulation is at the enzyme pyruvate kinase. This enzyme converts phosphoenolpyruvate into pyruvate. It is inhibited by ATP, acetyl-CoA, and long chain fatty acids, which are all markers of high energy levels.
In general, the overall regulation of glycolysis is related to how much energy the cell has. In energy rich states (ie: high ATP and low ADP/AMP) the cell slows glycolysis so that it can store glucose for use at a latter time. In energy depleted states (ie: low ATP and high ADP/AMP) the rate of glycolysis increases so that more energy can be formed by "burning" more glucose molecules per unit time.
Overview
Glycolysis breaks down glucose molecules. In the process the energy rich molecules ATP and NADH are formed. It is regulated at several enzymatic steps, most importantly at the enzyme phosphofructokinase-1. Pyruvate can be further metabolized to acetyl-CoA.
References and Resources
- Nelson DL, Cox MM. Lehninger Principles of Biochemistry. Fifth Edition. New York: Worth Publishers, 2008.
- Champe PC. Lippincott’s Illustrated Reviews: Biochemistry. Second Edition. Lippencott-Ravens Publishers, 1992.
- Le T, Bhushan V, Grimm L. First Aid for the USMLE Step 1. New York: McGraw Hill, 2009.